세포독성 T세포
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
세포독성 T세포는 CD8 분자를 발현하는 T 세포로, 바이러스 감염 세포나 종양 세포를 직접 파괴하는 역할을 하는 면역 세포이다. 흉선에서 발달하며, T 세포 수용체(TCR)를 통해 항원을 인식한다. 활성화된 세포독성 T세포는 퍼포린, 그랜자임 등을 분비하여 세포자멸사를 유도하거나, FasL을 통해 Fas 수용체를 자극하여 세포 사멸을 유도한다. 이들은 바이러스 감염, 류마티스 관절염, HIV 감염, 1형 당뇨병 등 다양한 질병의 발병 기전에 관여하며, 알레르기 반응에도 기여할 수 있다.

더 읽어볼만한 페이지
- T 세포 - CD4
CD4는 보조 T 세포 표면에서 T 세포 수용체의 공동수용체로 작용하여 항원제시세포와 상호작용을 돕고 T 세포 활성화 신호를 증폭시키는 면역계의 당단백질이며, HIV의 숙주 세포 침투 수용체로 작용하여 HIV 감염 및 AIDS 발병에 중요한 역할을 한다. - T 세포 - T세포 수용체
T세포 수용체(TCR)는 T 세포 표면의 이량체 단백질 복합체로, MHC 분자에 결합된 펩타이드를 인식하며, CD3 분자와 복합체를 이루어 신호 전달을 매개하고, V(D)J 재조합을 통해 다양한 항원 특이성을 갖는 T 세포 생성에 기여하여, CAR-T 치료 등 면역 요법 발전에 기여했다. - 백혈구 - 과립구
과립구는 골수에서 생성되어 식세포 작용과 독성 물질 방출을 통해 면역 반응에 기여하며, 호중구, 호산구, 호염기구, 비만 세포로 분류된다. - 백혈구 - 호중구
호중구는 혈액 내에서 가장 풍부한 백혈구로서 세균 감염에 대한 신속한 면역 반응에 중요한 역할을 하며, 식세포 작용, 탈과립, 호중구 세포외덫 형성 등의 항미생물 작용을 수행하고 염증 반응을 조절하며 사이토카인을 분비한다. - 세포 - 체세포
체세포는 다세포 생물의 몸을 이루는 세포로서 생식세포와 구별되며 불임성을 가지고, 종에 따라 생식세포계열과 분리되기도 하지만 체세포 배 발생 능력을 보이기도 하며, 유전 정보, 복제, 유전자 변형, 바이오뱅킹, 세포 노화 연구 등에 활용된다. - 세포 - 세포질
세포질은 핵을 제외한 세포 내부를 채우는 물질로, 세포질 기질, 세포소기관, 세포질 내포물로 구성되어 세포 형태 유지, 항상성 유지, 세포소기관 지탱, 물질대사 관련 물질 저장 등의 기능을 수행하며 액체 또는 젤 형태를 띤다.

2. 발달
면역계는 수백만 개의 잠재적인 항원을 인식해야 하지만, 인체의 유전자 수는 제한적이다. 따라서 골수 내 수백만 개의 백혈구 DNA 재조합을 통해 다양한 항원 수용체를 생성한다. 이 과정에서 인체 내 조직에 결합하는 자가 반응성 백혈구는 흉선에서 제거되며, 이 흉선의 발달과 활동에 요오드가 필수적이다.[2]
T 세포 수용체(TCR)는 주로 α, β 사슬로 구성되지만, γ, δ 사슬을 가진 TCR(감마 델타 T 세포)도 존재하며, 이들은 비단백질 항원을 인식하고 사이토카인과 케모카인을 생성하며, 다른 세포와 상호작용하여 세포 용해를 유발할 수 있다.[4][5] 조혈모 세포는 골수에서 흉선으로 이동하여 TCR 유전자 재조합을 거쳐 다양한 TCR을 생성한다. 기능적인 TCR을 가진 T 세포는 CD4와 CD8 공동 수용체를 모두 발현하는 이중 양성 T 세포(DP)가 된다.
이중 양성 T 세포는 흉선에서 자가 항원에 대한 양성 선택과 음성 선택을 거친다.
- '''양성 선택''': 자가 MHC의 존재하에 외부 항원에 결합하는 이중 양성 T 세포에서 발생하며, 제시된 항원과 관련된 MHC에 따라 CD4+ 또는 CD8+로 분화된다. 양성 선택은 자가 MHC 분자를 인식할 수 있는 TCR을 선택한다.
- '''음성 선택''': MHC가 제시한 자가 항원에 너무 강하게 결합하는 이중 양성 T 세포는 자가면역을 유발할 수 있으므로 세포사멸을 겪는다.
이러한 과정을 통해 MHC-자가 항원 복합체에 약하게 결합하는 T 세포만이 양성 선택되고, 살아남은 세포는 TCR이 인식하는 항원에 따라 단일 양성 T 세포(CD4+ 또는 CD8+)로 분화한다.
2. 1. 세포독성 T 세포로의 분화
MHC 클래스 I 제한 항원에 의해 활성화된 후, CD8+ T 세포는 성숙하여 '''세포독성 T 세포'''가 된다.[35] 미분화 T 세포는 헬퍼 T 세포에 필요한 CD4 분자와 킬러 T 세포에 필요한 CD8 분자를 모두 발현하고 있다(이중 양성, DP). 그러나 T 세포가 성숙해짐에 따라 CD4와 CD8 중 어느 한쪽만 발현하게 되며(단일 양성, SP),[35] 최종적으로 헬퍼 T 세포 또는 킬러 T 세포로 분화한다.세포독성 T 세포는 표면에 CD8 분자를 발현하는 T 세포로부터 분화하므로, "CD8 양성 T 세포" 또는 "CD8+T 세포"라고도 불린다.
3. 활성화
성숙한 T 세포는 항원과의 접촉 횟수에 따라 순수 T 세포, 기억 T 세포, 이펙터 T 세포로 구분된다.[6][9] 순수 T 림프구는 가슴샘에 위치하며 항원에 아직 노출되지 않은 T 세포의 초기 단계이다. 기억 T 세포는 항원과 한 번 이상 접촉했지만 이후 휴지 상태로 돌아간 T 세포로, 특정 항원에 다시 반응할 준비가 된 상태이다. 이펙터 T 세포는 면역 반응이 유발되면 활성화되어 병원체나 종양 세포를 죽일 수 있는 세포이다.[6][9]
세포 활성화는 흉선 비의존적 경로(감염된 APC) 또는 흉선 의존적 경로(CD4+ T 세포)를 통해 일어난다. 흉선 비의존적 경로에서는 감염된 APC가 다수의 공동 수용체를 발현하여 활성화된다. 흉선 의존적 경로에서는 CD4+ T 세포가 APC를 활성화하거나 IL-2를 분비하여 세포독성 T 세포를 직접 활성화한다.
활성화된 세포독성 T 세포는 표적 세포에 "치명적인 타격"을 가하고, 표적 세포는 약 6시간 안에 세포 사멸을 통해 죽는다.[7]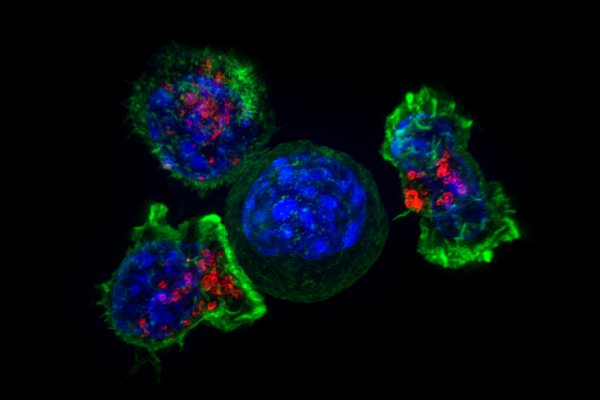
I형 MHC는 적혈구를 제외한 세포 핵이 있는 숙주 세포에 의해 발현된다. 세포가 세포내 병원체에 감염되면 세포는 항원 처리를 통해 외래 단백질을 분해하고, 생성된 펩타이드 조각 중 일부는 CD8+ T 세포의 T 세포 항원 수용체(TCR)에 의해 I형 MHC에 제시된다.
대부분 활성화는 항원의 TCR 인식을 통해 이루어지지만, 다른 경로를 통해서도 활성화가 가능하다. 예를 들어, 세포독성 T 세포는 다른 CD8 T 세포에 의해 표적이 되면 활성화되어 후자의 관용을 유도한다.[12]
3. 1. 활성화 과정
세포독성 T 세포(CTL)의 활성화는 T 세포 표면에 있는 분자와 항원 제시 세포(APC) 표면에 있는 분자 간의 여러 상호 작용을 통해 이루어진다. TC 세포 활성화를 위한 '''''두 신호 모델'''''은 다음과 같다.
순수한 CD8+ T 세포를 활성화하려면 주로 성숙된 수지상 세포를 포함한 전문적인 항원 제시 세포와의 상호 작용이 필요하다.[8] 장기간 지속되는 기억 T 세포를 생성하고 세포독성 T 세포의 반복적인 자극을 허용하기 위해, 수지상 세포는 활성화된 CD4+ 헬퍼 T 세포와 CD8+ T 세포 모두와 상호 작용해야 한다.[9] 이 과정에서 CD4+ 헬퍼 T 세포는 수지상 세포가 순수한 CD8+ T 세포에 강력한 활성화 신호를 제공하도록 "허가"한다.[10] CD4+ T 헬퍼 세포에 의한 항원 제시 세포의 이러한 허가는 면역 시냅스 형성 동안 T 헬퍼 세포의 CD154/CD40L과 항원 제시 세포의 CD40 수용체 간의 신호를 통해 진행된다.[11]
헬퍼 T 세포(Th) 중 Th1 세포는 주로 인터류킨-2(IL-2) 및 IFN-γ를 생성하여 세포독성 T 세포의 작용을 보조한다.
일단 활성화되면 TC 세포는 T 세포의 성장 및 분화 인자인 인터류킨 2 (IL-2)의 도움을 받아 클론 확장을 겪는다.
4. 작동 기전 (Effector functions)
활성화된 TC 세포는 감염되었거나 기능 이상이 있는 체세포를 만나면 퍼포린, 그랜자임, 과립용해소와 같은 세포독성 물질을 방출한다. 퍼포린은 표적 세포막에 구멍을 뚫어 그랜자임이 세포질 안으로 들어가게 한다. 세린 프로테아제인 그랜자임은 카스파제 연쇄반응을 일으켜 세포자멸사를 유도한다. 이러한 표적 세포의 파도와 같은 죽음을 "치명타"라고 부른다.[13] TC 세포는 세포막에 높은 지질 정렬과 음전하를 띤 포스파티딜세린이 있어 퍼포린과 그랜자임의 영향에 저항성을 가진다.[14]
세포자멸사를 유도하는 또 다른 방법은 TC 세포와 감염된 세포 간의 세포 표면 상호작용이다. 활성화된 TC 세포는 표면에 FAS 리간드(FasL)를 발현하며, 이는 표적 세포의 Fas와 결합한다. 이 결합은 세포사멸 유도 신호 복합체(DISC)를 형성하게 한다.[15] Fas 연관 사멸 도메인(FADD)은 DISC와 함께 프로카스파제 8과 10을 모으고,[15] 이 카스파제들은 이펙터 카스파제 3, 6, 7을 활성화시켜 라민 A, 라민 B1, 라민 B2, 폴리 ADP 리보스 중합효소(PARP), DNA-PKcs와 같은 사멸 기질을 절단한다. 최종적으로 Fas를 발현하는 세포는 세포자멸사로 이어진다. CD8 T 세포는 CD3 수용체 복합체에 의해 매개되는 활성화 유도 세포 사멸(AICD)을 보일 수도 있다. 최근 혈소판에서 방출된 단백질 TLT-1이 CD8 T 세포에서 AICD 유사 세포 사멸을 유도하는 것으로 밝혀졌다.[16]
전사 인자 Eomesodermin은 적응 면역 반응에서 조절 유전자 역할을 하며, CD8+ T 세포 기능에 중요한 역할을 한다.[17] Eomesodermin의 기능 상실 영향을 조사한 연구에서 이 전사 인자의 발현 감소는 CD8+ T 세포에서 생성되는 퍼포린의 양을 감소시키는 것으로 나타났다.[17]
5. 질병에서의 역할 (Role in disease pathogenesis)
B형 간염 바이러스(HBV) 감염에서 세포독성 T세포는 감염된 세포를 제거하고 항바이러스 사이토카인을 생성하여 HBV를 제거하지만, 동시에 간 손상의 주요 원인이 된다.[19] 혈소판은 바이러스 특이적 세포독성 T세포가 감염된 간에 축적되도록 돕는다.[20] 쥐 실험에서 CXCR5+CD8+T 세포 주입은 HBsAg를 감소시켰고, CXCL13 수치 증가는 간내 CXCR5+CD8+T 세포 모집을 촉진하여 만성 HBV 감염 관리에 도움을 줄 수 있는 높은 수준의 HBV 특이적 인터페론(IFN)-γ 및 IL-21을 생성했다.[21]
세포독성 T세포는 관절염 진행, 특히 류마티스 관절염에서 관절 염증에 관여한다. 활막은 과형성, 혈관 증가, 염증 세포 침윤이 특징이며, CD4+ T 림프구가 세포 매개 면역 반응의 주요 조절자이다. 류마티스 관절염은 주요 조직 적합성 복합체(MHC) 클래스 II 항원과 관련이 있으며, 이는 류마티스 관절염이 외부 항원(바이러스 단백질 등) 또는 내인성 단백질 항원에 의해 발생함을 시사한다.[22] 활성화된 CD4+ T 림프구는 단핵구, 대식세포, 활막 섬유아세포를 자극하여 인터류킨-1, 인터류킨-6, 종양 괴사 인자 알파(TNFα)를 생성하고, B 세포를 자극하여 류마티스 인자를 포함한 면역글로불린을 생성한다.[23] 세포독성 T세포의 사이토카인 생성은 관절염 질환 진행을 가속화할 수 있다.[24]
CD8+ T 세포는 HIV 감염에서 중요한 역할을 한다. HIV는 높은 돌연변이율과 감염 세포 표면 MHC 클래스 I 단백질 발현 하향 조절을 통해 CD8+ T 세포의 인식을 회피한다.[25]
CD8+ T 세포는 1형 당뇨병에서 이자 베타 세포를 파괴하는 주요 이펙터 세포이다.[26] 베타-2 마이크로글로불린(B2M) 유전자좌에 돌연변이가 있어 MHC 클래스 I 분자와 CD8+ T 세포가 없는 NOD 생쥐는 당뇨병이 발생하지 않는다.[27]
CD8+ T 세포는 화학요법 유발 말초 신경병증(CIPN) 해결에 필요할 수 있다.[28][29]
세포독성 T 림프구는 이식 거부 반응,[30] 심각한 SARS-CoV-2 감염 시 과도한 사이토카인 생성,[31][32] 다발성 경화증과 같은 중추 신경계 염증 및 퇴행성 질환에도 관여한다.[33]
6. 알레르기
세포독성 T 세포가 과도하게 반응하여 알레르기를 유발하는 경우가 있다. 그 유발 요인(방아쇠)은 개인의 체질에 따른다.
7. 임상 응용
임상적으로, 암에 대해 암세포 특이적인 항원에 대한 세포독성 T 세포(CTL)를 유도함으로써 치료하려는 면역 요법 등이 연구되고 있다.[36]
참조
[1]
서적
Advanced Hematology in Integrated Cardiovascular Chinese Medicine
Elsevier
[2]
논문
Iodine, thymus, and immunity
2009-09
[3]
논문
Features and functions of gamma delta T lymphocytes: focus on chemokines and their receptors
2003
[4]
논문
Ligand recognition by the γδ TCR and discrimination between homeostasis and stress conditions
2020-09
[5]
논문
Characterization of Adaptive-like γδ T Cells in Ugandan Infants during Primary Cytomegalovirus Infection
2021-10
[6]
서적
Inmunología (de memoria)
https://www.worldcat[...]
Médica Panamericana
2017
[7]
서적
Cellular and molecular immunology
https://www.worldcat[...]
2018
[8]
논문
Crosstalk between T lymphocytes and dendritic cells
2012
[9]
논문
Concurrent interaction of DCs with CD4(+) and CD8(+) T cells improves secondary CTL expansion: It takes three to tango
2014-12
[10]
논문
Immunology. Licence to kill
1998-06
[11]
논문
Help for cytotoxic-T-cell responses is mediated by CD40 signalling
1998-06
[12]
논문
CTLs respond with activation and granule secretion when serving as targets for T-cell recognition
2011-01
[13]
논문
Preparing the lethal hit: interplay between exo- and endocytic pathways in cytotoxic T lymphocytes
2017-02
[14]
논문
Lipid order and charge protect killer T cells from accidental death
2019-11
[15]
서적
Encyclopedia of Medical Immunology
[16]
논문
Platelet-derived TLT-1 promotes tumor progression by suppressing CD8+ T cells
[17]
논문
Control of effector CD8+ T cell function by the transcription factor Eomesodermin
2003-11
[18]
논문
Type B coxsackieviruses and their interactions with the innate and adaptive immune systems
2010-09
[19]
논문
Pathogenetic and antiviral immune responses against hepatitis B virus
[20]
논문
Platelets mediate cytotoxic T lymphocyte-induced liver damage
2005-11
[21]
논문
CXCL13-mediated recruitment of intrahepatic CXCR5+CD8+ T cells favors viral control in chronic HBV infection
2020-03
[22]
논문
Antibody-dependent and -independent mechanisms of inflammatory arthritis
2019-03
[23]
논문
The central role of T cells in rheumatoid arthritis
https://pubmed.ncbi.[...]
2007-09
[24]
논문
Potential roles for CD8(+) T cells in rheumatoid arthritis
2013-01
[25]
논문
CD8+ T-cells: function and response to HIV infection
2004-01
[26]
서적
Immunopathogenesis of Type 1 Diabetes Mellitus
2008
[27]
논문
The role of CD8+ T cells in the initiation of insulin-dependent diabetes mellitus
1996-08
[28]
논문
Cisplatin educates CD8+ T cells to prevent and resolve chemotherapy-induced peripheral neuropathy in mice
2019-06
[29]
논문
CD8+ T Cells and Endogenous IL-10 Are Required for Resolution of Chemotherapy-Induced Neuropathic Pain
2016-10
[30]
논문
Human leukocyte antigen molecular mismatch to risk stratify kidney transplant recipients
2020-02
[31]
논문
Pathogenic human coronavirus infections: causes and consequences of cytokine storm and immunopathology
2017-07
[32]
논문
COVID-19, cytokines and immunosuppression: what can we learn from severe acute respiratory syndrome?
2020-03
[33]
논문
Cytotoxic T lymphocytes in autoimmune and degenerative CNS diseases
2002-06
[34]
서적
宮坂昌之 ほか編集『標準免疫学』
医学書院
2016-02-01
[35]
서적
宮坂昌之 ほか編集『標準免疫学』
医学書院
2016-02-01
[36]
서적
エッセンシャル免疫学
メディカル・サイエンス・インターナショナル
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com